Research progress on dark fermentative bio-hydrogen production from lignocellulose assisted by metal nanoparticles
TONG Haihang,, SHI Dezhi,, LIU Jiayu, CAI Huayi, LUO Dan, CHEN Fei
College of Environment and Ecology, Key Laboratory of the Three Gorges Reservoir Region’s Eco-Environment, Ministry of Education, Chongqing University, Chongqing 400045, China
Based on bibliometric analysis of bio-hydrogen published 2004 to 2020 indexed by Web of Science, dark fermentation as one of the four methods of bio-hydrogen, has received the highest attention for its assets such as high hydrogen-production rate without light, low energy consumption and comprehensively available microorganism and substrate. At the same time, some metal nanoparticles (Fe, Ni, Co, and Ag) that exhibit excellent promotion for dark fermentation has been spotlight in this research field. Nanoparticles equipped with the properties and characteristics of small volume, surface effect and quantum size effect can enhance the rate of electron transfer of cells and present good biocompatibility. The mechanism of metal nanoparticles (MNPs) interacting with enzymes and microoganisms has been illustrated. Particularly, impacts of MNPs are elaborated in details on different mechanisms of enhancing the hydrogen production process through dark fermentation by promoting hydrolysis of lignocellulose, immobilization of cellulase enzymes, and activity of hydrogenase, respectively. On one hand, MNPs assist pretreatment process of removing of lignin, and improve stability of enzymes by immobilizing them; on the other hand, MNPs enhance the rate of electron transfer, thus make a promotion of activity of hydrogenase, and finally lead the increase of hydrogen production. In addition to the impact of nanoparticles on enzymes, the role of nanoparticles on bacterial metabolism, interspecies electron transfer and synergistic hydrogen production of mixed culture are discussed. The difficulties and prospects of the application of metal nanoparticles in dark fermentation for hydrogen production are also prospected.
Keywords:lignocellulose
;
dark fermentative
;
bio-hydrogen
;
metal nanoparticles
;
enzyme
;
microorganism
TONG Haihang, SHI Dezhi, LIU Jiayu, CAI Huayi, LUO Dan, CHEN Fei. Research progress on dark fermentative bio-hydrogen production from lignocellulose assisted by metal nanoparticles[J]. CIESC Journal, 2022, 73(4): 1417-1435. doi:10.11949/0438-1157.20211412
生物制氢主要有四种途径:光解水产氢、光发酵产氢、暗发酵产氢和光-暗联合发酵产氢[6-9]。暗发酵由于无须光照、产氢速率快、耗能低、微生物类群广泛且可使用多种有机底物,是生物制氢最常用的工艺[10-12]。Web of Science引文索引中2004~2020年间涉及生物制氢各种主要方法的文章发表数量如图1(a)所示。可知暗发酵产氢相关的文章数量明显高于其他几种方法,且随年份呈现迅速增长趋势,近五年来尤为显著,2020年发表的文章数量接近250篇。相比之下其他几种生物制氢方法的文章数量较少且随年份增长缓慢。暗发酵产氢主要通过三种途径:丁酸型发酵、混合酸发酵以及NADH途径。参与暗发酵的碳水化合物主要是葡萄糖,主要产生乙酸、丁酸与氢气,具体反应式如式(1)~式(4):
金属纳米颗粒通过提高氢化酶的活性促进暗发酵产氢过程已经得到界内的认可,一些学者近年来利用铁、镍和其他金属纳米颗粒及其掺杂复合材料等来促进暗发酵产氢的研究总结见表2。Han等[60]研究了不同浓度的Fe2O3NPs对以丁酸梭菌为优势菌群的混合厌氧细菌培养系统在以蔗糖为底物的间歇反应器中介导的暗发酵生物制氢的影响,发现在最适温度35℃,pH=8.48,纳米颗粒的浓度为200 mg/L的条件下,最大产氢量达3.21 mol H2/(mol蔗糖),比对照组(添加量为0)的产氢量高32.64%。不同金属纳米颗粒的复合材料也有相应的研究,Beckers等[110]研究了包裹在多孔SiO2中的Fe x O y 、Pd、Ag、Cu复合纳米颗粒(FeNPs@SiO2,PdNPs@SiO2,AgNPs@SiO2,CuNPs@SiO2)对暗发酵厌氧丁酸梭菌产氢的影响,发现与对照组(不添加纳米颗粒)相比,复合纳米颗粒均对产氢有促进作用,其中FeNPs@SiO2纳米颗粒的作用最明显,氢气总产量提高了38%,平均比产氢速率从44.7 ml/h提高到61.8 ml/h。
Table 2
表2
表2不同类型金属纳米材料在暗发酵生物制氢中的应用
Table 2 Application of different MNPs in dark fermentative hydrogen production
Fig.7
Relation between addition of MNPs and H2 production in dark fermentation from lignocellulose[91, 95, 102-104, 115]
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量。Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性。Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果。37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%。这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度。
A novel strategy to enhance biohydrogen production using graphene oxide treated thermostable crude cellulase and sugarcane bagasse hydrolyzate under co-culture system
Overcoming challenges in lignocellulosic biomass pretreatment for second-generation (2G) sugar production: emerging role of nano, biotechnological and promising approaches
Effect of nickel-cobaltite nanoparticles on production and thermostability of cellulases from newly isolated thermotolerant Aspergillus fumigatus NS (class: Eurotiomycetes)
[J]. Applied Biochemistry and Biotechnology, 2014, 174(3): 1092-1103.
Synthesis of magnetic gold mesoporous silica nanoparticles core shell for cellulase enzyme immobilization: improvement of enzymatic activity and thermal stability
Immobilization of cellulase enzyme on zinc ferrite nanoparticles in increasing enzymatic hydrolysis on ultrasound-assisted alkaline pretreated crotalaria juncea biomass
[J]. Indian Journal of Science and Technology, 2017, 10(24): 1-7.
Nanoparticle-associated single step hydrogen fermentation for the conversion of starch potato waste biomass by thermophilic Parageobacillusthermoglucosidasius
Explore the possible effect of TiO2 and magnetic hematite nanoparticle addition on biohydrogen production by Clostridium pasteurianum based on gene expression measurements
[J]. International Journal of Hydrogen Energy, 2016, 41(46): 21685-21691.
Enhanced fermentative hydrogen production from industrial wastewater using mixed culture bacteria incorporated with iron, nickel, and zinc-based nanoparticles
Comparative evaluation of fermentative hydrogen production using Enterobacter cloacae and mixed culture: effect of Pd (Ⅱ) ion and phytogenic palladium nanoparticles
Role of L-cysteine and iron oxide nanoparticle in affecting hydrogen yield potential and electronic distribution in biohydrogen production from dark fermentation effluents by photo-fermentation
[J]. Journal of Cleaner Production, 2020, 276: 123193.
Revealing the mechanisms of alkali-based magnetic nanosheets enhanced hydrogen production from dark fermentation: comparison between mesophilic and thermophilic conditions
Improving hydrogen and methane co-generation in cascading dark fermentation and anaerobic digestion: the effect of magnetite nanoparticles on microbial electron transfer and syntrophism
[J]. Chemical Engineering Journal, 2020, 397: 125394.
A new model for electron flow during anaerobic digestion: direct interspecies electron transfer to Methanosaeta for the reduction of carbon dioxide to methane
[J]. Energy and Environmental Science, 2014, 7(1): 408-415.
High yield expression of an AHL-lactonase from Bacillus sp. B546 in Pichia pastoris and its application to reduce Aeromonas hydrophila mortality in aquaculture
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
A novel strategy to enhance biohydrogen production using graphene oxide treated thermostable crude cellulase and sugarcane bagasse hydrolyzate under co-culture system
Beyond the theoretical yields of dark-fermentative biohydrogen
3
2018
... 生物制氢主要有四种途径:光解水产氢、光发酵产氢、暗发酵产氢和光-暗联合发酵产氢[6-9].暗发酵由于无须光照、产氢速率快、耗能低、微生物类群广泛且可使用多种有机底物,是生物制氢最常用的工艺[10-12].Web of Science引文索引中2004~2020年间涉及生物制氢各种主要方法的文章发表数量如图1(a)所示.可知暗发酵产氢相关的文章数量明显高于其他几种方法,且随年份呈现迅速增长趋势,近五年来尤为显著,2020年发表的文章数量接近250篇.相比之下其他几种生物制氢方法的文章数量较少且随年份增长缓慢.暗发酵产氢主要通过三种途径:丁酸型发酵、混合酸发酵以及NADH途径.参与暗发酵的碳水化合物主要是葡萄糖,主要产生乙酸、丁酸与氢气,具体反应式如式(1)~式(4): ...
Advances in nanomaterials induced biohydrogen production using waste biomass
7
2020
... 生物制氢主要有四种途径:光解水产氢、光发酵产氢、暗发酵产氢和光-暗联合发酵产氢[6-9].暗发酵由于无须光照、产氢速率快、耗能低、微生物类群广泛且可使用多种有机底物,是生物制氢最常用的工艺[10-12].Web of Science引文索引中2004~2020年间涉及生物制氢各种主要方法的文章发表数量如图1(a)所示.可知暗发酵产氢相关的文章数量明显高于其他几种方法,且随年份呈现迅速增长趋势,近五年来尤为显著,2020年发表的文章数量接近250篇.相比之下其他几种生物制氢方法的文章数量较少且随年份增长缓慢.暗发酵产氢主要通过三种途径:丁酸型发酵、混合酸发酵以及NADH途径.参与暗发酵的碳水化合物主要是葡萄糖,主要产生乙酸、丁酸与氢气,具体反应式如式(1)~式(4): ...
... 生物制氢主要有四种途径:光解水产氢、光发酵产氢、暗发酵产氢和光-暗联合发酵产氢[6-9].暗发酵由于无须光照、产氢速率快、耗能低、微生物类群广泛且可使用多种有机底物,是生物制氢最常用的工艺[10-12].Web of Science引文索引中2004~2020年间涉及生物制氢各种主要方法的文章发表数量如图1(a)所示.可知暗发酵产氢相关的文章数量明显高于其他几种方法,且随年份呈现迅速增长趋势,近五年来尤为显著,2020年发表的文章数量接近250篇.相比之下其他几种生物制氢方法的文章数量较少且随年份增长缓慢.暗发酵产氢主要通过三种途径:丁酸型发酵、混合酸发酵以及NADH途径.参与暗发酵的碳水化合物主要是葡萄糖,主要产生乙酸、丁酸与氢气,具体反应式如式(1)~式(4): ...
A comparative overview of hydrogen production processes
0
2017
Biohydrogen production from de-oiled rice bran as sustainable feedstock in fermentative process
1
2016
... 生物制氢主要有四种途径:光解水产氢、光发酵产氢、暗发酵产氢和光-暗联合发酵产氢[6-9].暗发酵由于无须光照、产氢速率快、耗能低、微生物类群广泛且可使用多种有机底物,是生物制氢最常用的工艺[10-12].Web of Science引文索引中2004~2020年间涉及生物制氢各种主要方法的文章发表数量如图1(a)所示.可知暗发酵产氢相关的文章数量明显高于其他几种方法,且随年份呈现迅速增长趋势,近五年来尤为显著,2020年发表的文章数量接近250篇.相比之下其他几种生物制氢方法的文章数量较少且随年份增长缓慢.暗发酵产氢主要通过三种途径:丁酸型发酵、混合酸发酵以及NADH途径.参与暗发酵的碳水化合物主要是葡萄糖,主要产生乙酸、丁酸与氢气,具体反应式如式(1)~式(4): ...
CFD optimization of continuous stirred-tank (CSTR) reactor for biohydrogen production
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
... [24]SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
... [26, 88]Chemical structure of the active site for hydrogenase<sup>[<xref ref-type="bibr" rid="R26">26</xref>, <xref ref-type="bibr" rid="R88">88</xref>]</sup>Fig.52Fe0+O2+2H2O2Fe2++4OH-, ΔE0=1.67 V
... [35]Interaction between nanoparticles and cell(receptor mediated endocytosis: nanoparticles are coated by a protein corona)<sup>[<xref ref-type="bibr" rid="R35">35</xref>]</sup>Fig.21.2 金属纳米颗粒与酶及微生物的作用机理
... [43]Enzyme stabilization by immobilization introduces additional covalent and non-covalent forces to nanoparticles<sup>[<xref ref-type="bibr" rid="R43">43</xref>]</sup>Fig.3
... [49-50]Hydrolysis of lignocellulose by functionalized magnetic nanoparticles<sup>[<xref ref-type="bibr" rid="R49">49</xref>-<xref ref-type="bibr" rid="R50">50</xref>]</sup>Fig.42.1.2 金属纳米颗粒对酶的固定化作用强化酶对木质纤维素的预处理
... -50]Hydrolysis of lignocellulose by functionalized magnetic nanoparticles<sup>[<xref ref-type="bibr" rid="R49">49</xref>-<xref ref-type="bibr" rid="R50">50</xref>]</sup>Fig.42.1.2 金属纳米颗粒对酶的固定化作用强化酶对木质纤维素的预处理
Overcoming challenges in lignocellulosic biomass pretreatment for second-generation (2G) sugar production: emerging role of nano, biotechnological and promising approaches
... 金属纳米颗粒通过提高氢化酶的活性促进暗发酵产氢过程已经得到界内的认可,一些学者近年来利用铁、镍和其他金属纳米颗粒及其掺杂复合材料等来促进暗发酵产氢的研究总结见表2.Han等[60]研究了不同浓度的Fe2O3NPs对以丁酸梭菌为优势菌群的混合厌氧细菌培养系统在以蔗糖为底物的间歇反应器中介导的暗发酵生物制氢的影响,发现在最适温度35℃,pH=8.48,纳米颗粒的浓度为200 mg/L的条件下,最大产氢量达3.21 mol H2/(mol蔗糖),比对照组(添加量为0)的产氢量高32.64%.不同金属纳米颗粒的复合材料也有相应的研究,Beckers等[110]研究了包裹在多孔SiO2中的Fe x O y 、Pd、Ag、Cu复合纳米颗粒(FeNPs@SiO2,PdNPs@SiO2,AgNPs@SiO2,CuNPs@SiO2)对暗发酵厌氧丁酸梭菌产氢的影响,发现与对照组(不添加纳米颗粒)相比,复合纳米颗粒均对产氢有促进作用,其中FeNPs@SiO2纳米颗粒的作用最明显,氢气总产量提高了38%,平均比产氢速率从44.7 ml/h提高到61.8 ml/h. ...
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
Application of magnetic nanoparticles in smart enzyme immobilization
1
2016
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
Encapsulation of omega-3 fatty acids in nanoemulsions and microgels: Impact of delivery system type and protein addition on gastrointestinal fate
1
2017
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
Protein-nanoparticle interactions
1
2008
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
Advances in enzyme immobilisation
1
2009
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
Immobilization of cellulase onto core-shell magnetic gold nanoparticles functionalized by aspartic acid and determination of its activity
2
2020
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Characterization of direct cellulase immobilization with superparamagnetic nanoparticles
2
2011
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Immobilization of cellulase on thermo-sensitive magnetic microspheres: improved stability and reproducibility
1
2018
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Molecular imprinting and immobilization of cellulase onto magnetic Fe3O4@SiO2 nanoparticles
1
2014
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Immobilization of laccase on magnetic silica nanoparticles and its application in the oxidation of guaiacol, a phenolic lignin model compound
2
2015
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
... 此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Application of molecular imprinted magnetic Fe3O4@SiO2 nanoparticles for selective immobilization of cellulase
2
2016
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
... 此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Cellulases immobilization on chitosan-coated magnetic nanoparticles: application for agave atrovirens lignocellulosic biomass hydrolysis
1
2017
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Preparation of magnetic chitosan nanoparticles as support for cellulase immobilization
1
2014
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Metal affinity immobilization of cellulase on Fe3O4 nanoparticles with copper as ligand for biocatalytic applications
2
2019
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
... 此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Innovative hydrolysis of corn stover biowaste by modified magnetite laccase immobilized nanoparticles
2
2020
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Immobilization of cellulases on amine and aldehyde functionalized Fe2O3 magnetic nanoparticles
1
2012
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Immobilization of cellulase on functionalized cobalt ferrite nanoparticles
1
2016
... Application of MNPs on cellulose or laccase immobilizationTable 1
序号
纳米材料
固定/结合模式
条件参数
1
Fe3O4 NPs [68]
戊二醛表面修饰共价结合
T=60℃, pH=4.5
2
Fe3O4 NPs [69]
通过半胱氨酸基团的表面功能化卵清蛋白结合
T =80℃, pH=4.5
3
Fe3O4@SiO2[70]
物理吸附
T =70℃, pH=4.0
4
Fe3O4 NPs @SiO2[71]
APTES共价结合
T =25℃, pH=4
5
Fe3O4 NPs @SiO2[72]
甲基丙烯酸缩水甘油酯表面功能化共价结合
T =50℃, pH=5.0
6
Fe3O4@SiO2@KIT-6NPs[54]
APTES共价结合
T =35℃, pH=4.5
7
Fe3O4 NPs /壳聚糖[73]
戊二醛表面修饰共价结合
T =60℃, pH=5.5
8
Fe3O4 NPs/壳聚糖[74]
戊二醛表面修饰共价结合
T =60℃, pH=5.0
9
Cu/Fe3O4 NPs[75]
APTES共价结合
T =80℃, pH=5
10
Cu2+ modified Fe3O4-NH2NPs[76]
亲和吸附
T =35℃, pH=4.5
11
Fe3O4@Au NPs[67]
通过聚乙二醇和L-天门冬氨酸共价结合
T =50℃, pH=4.8
12
Fe2O3 NPs[77]
戊二醛表面修饰共价结合
T =50℃, pH=4.8
13
CoFe2O4 NPs [78]
基于EDS & NHS的戊二醛结合
T =50℃, pH=5.0
14
MnO2 NPs [46]
戊二醛表面改性卵圆结合
T =70℃, pH=5.0
此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Effect of nickel-cobaltite nanoparticles on production and thermostability of cellulases from newly isolated thermotolerant Aspergillus fumigatus NS (class: Eurotiomycetes)
1
2014
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
Effect of cell immobilization, hematite nanoparticles and formation of hydrogen-producing granules on biohydrogen production from sucrose wastewater
2
2017
... 纳米颗粒通过共价结合或吸附固定在蛋白质或酶上(如图3),对酶的催化活性产生很大影响.酶被附着并固定在纳米颗粒表面上,形成纳米颗粒-蛋白质冠(nanoparticle-protein corona),这种蛋白质冠可导致酶系统的某些结构和功能发生变化,从而增强纳米颗粒的生物相容性[63].在纳米颗粒-蛋白质冠形成过程中,蛋白质在纳米颗粒上的排列模式进一步影响纳米颗粒的生物活性[64].酶蛋白对纳米颗粒的亲和力及其占据纳米颗粒表面的能力在酶的固定化作用中起着决定性的作用[65].酶可以通过范德华力、静电力、疏水或π-π键堆积连接等非共价结合方式与金属纳米颗粒结合.然而,非共价结合存在蛋白质从金属纳米颗粒表面脱落的问题.通过使用交联剂(如戊二醛)实现酶与金属纳米颗粒的共价结合是使用最广泛的酶固定化方法.在共价固定化技术中,纳米颗粒载体表面的化学基团被激活,并与酶蛋白质的赖氨酸氨基侧链、半胱氨酸的巯基、天冬氨酸和谷氨酸残基的咪唑和酚基等发生特异性反应进行结合,形成具有较高强度和稳定性的共价键[66-67].表1总结了一些学者利用金属纳米颗粒进行纤维素酶和漆酶固定化的研究.众多研究表明采用不同纳米颗粒处理纤维素酶后,纤维素酶的稳定性均得到提高.Cherian等[46]研究了金属纳米颗粒对蛋白质稳定性的影响,发现固定在MnO2NPs上的酶在25℃下的热稳定时间比对照组增加了2 h.Xu等[68]将纤维素酶固定在戊二醛官能化的Fe3O4NPS上,发现其比游离酶具有更好的热稳定性和酸碱稳定性.固定化有助于提高酶对纤维素底物的亲和力,因此可以获得更好的水解性能.Srivastava等[79]用NiCo2O4NPs处理纤维素酶,与对照组在80℃下测试0~8 h,结果表明,纤维素酶在2 h和4 h内的相对活性分别保持在63%和50%,7 h后相对活性比开始时下降了21%.与对照组相比,纤维素酶的稳定性分别提高了68%、50%和41%.Salem等[80]将从废弃污泥中获得的产氢细菌细胞固定在Fe2O3NPs上,使比产氢速率由3.87 L/(L·d) 增加至5.9 L/(L·d),H2产量由0.19 L H2/(g蔗糖)增加至0.3 L H2/(g蔗糖).Ramírez[61]发现相较于游离纤维素酶(最适温度为50℃,最适pH范围为4.5~5.0),将纤维素酶固定在壳聚糖包覆的Fe3O4NPs上,催化性能提高了1.5倍,热稳定性提高至60℃,pH提高至5.5.且在60℃和70℃孵育4 h后,经稳定的纤维素酶的储存稳定性为初始活性的50%,而未经稳定的纤维素酶的活性仅为初始活性的20%.木质素的降解主要依靠漆酶,Amin等[54]将漆酶固定在改性的Fe3O4@SiO2@KIT-6 NPs上,可以增强漆酶对橄榄果渣的脱木质素作用,在35℃的最佳温度下木质素的去除率高达77.3%.Gou等[76]将漆酶固定在Cu2+改性的Fe3O4-NH2纳米颗粒上进行木质纤维素的预处理,结果显示处理后玉米芯中的木质素降解率达40.76%,在随后的纤维素水解过程中纤维素转化率达到38.37%,同对照组相比提高了23.98%. ...
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Synthesis of magnetic gold mesoporous silica nanoparticles core shell for cellulase enzyme immobilization: improvement of enzymatic activity and thermal stability
1
2018
... 此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Immobilization of cellulase enzyme on zinc ferrite nanoparticles in increasing enzymatic hydrolysis on ultrasound-assisted alkaline pretreated crotalaria juncea biomass
1
2017
... 此外,纳米颗粒对酶的稳定化还体现在提高酶的重复利用性和储存稳定性上.Tao等[72]利用Fe3O4NPs@SiO2固定纤维素酶,发现在50℃条件下,经固定的纤维素酶的半衰期是未经处理的纤维素酶的2倍,且在重复使用7个循环后,依然能保持最初活性的77%.Poorakbar等[81]制备了一种磁性介孔SiO2纳米颗粒用于纤维素酶的固定,经固定的纤维素酶在9 h后的活性能保持初始活性58%,热稳定性、pH稳定性和储存稳定性均得到提升.Abbaszadeh等[75]用铜负载的磁铁矿纳米颗粒稳定纤维素酶,发现经稳定后的纤维素酶展现出更好的稳定性,并且在循环使用5次后其活性依然能保持初始活性的73%,在4℃下保存8 d后,未经稳定和经稳定的纤维素酶的活性分别为初始活性的70%和84%.Manasa等[82]利用铁酸锌纳米颗粒(Zn x Fe3-x O4NPs)固定纤维素酶,使酶的活性增加,在同样的超声波辅助碱性预处理条件下纤维素酶水解的糖产量增加53%,并且固定化纤维素酶的活性在60℃下保持了3个循环.Hu等[71]制备了一种利用氨基功能化磁性二氧化硅纳米颗粒并研究其对漆酶的稳定化效果,经稳定化的漆酶在50℃下孵育2 h后活性能保持初始活性的98%,而未经处理的漆酶活性仅为初始活性的40%.将未经稳定和稳定后的漆酶在4℃下储存,18 d后未经稳定的漆酶完全失活,而稳定后的漆酶活性仍能保持初始活性的79.25%. ...
Peer reviewed: extracting hydrogen and electricity from renewable resources
... , 88]Chemical structure of the active site for hydrogenase<sup>[<xref ref-type="bibr" rid="R26">26</xref>, <xref ref-type="bibr" rid="R88">88</xref>]</sup>Fig.52Fe0+O2+2H2O2Fe2++4OH-, ΔE0=1.67 V
Relation between addition of MNPs and H<sub>2</sub> production in dark fermentation from lignocellulose<sup>[<xref ref-type="bibr" rid="R91">91</xref>, <xref ref-type="bibr" rid="R95">95</xref>, <xref ref-type="bibr" rid="R102">102</xref>-<xref ref-type="bibr" rid="R104">104</xref>, <xref ref-type="bibr" rid="R115">115</xref>]</sup>Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... [91, 95, 102-104, 115]Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... 目前针对暗发酵产氢过程中微生物群落变化的研究主要是通过高通量测序观察微生物的相对丰度变化.Yang等[99]研究了铁纳米颗粒添加对暗发酵微生物群落的影响,发现相较于对照组,添加Fe0NPS、carbon/Fe0NPs样品的Simpson指数显著降低,Shannon指数显著升高,表明这两种添加剂显著提高了系统中微生物的多样性.此外,还可使肠杆菌属(Enterobacter sp.)和梭菌属(Clostridium sp.)的相对丰度存在显著差异.对照组、Fe0NPS和carbon/Fe0NPs的实验组中Enterobacter sp.分别占87.8%、52.8%和15.6%,Clostridium sp.分别占7.2%、40.3%和77.5%.Zhang等[100]研究了零价铁粉末-活性炭(ZVI-AC)强化生物制氢过程中微生物群落的变化,发现添加了ZVI-AC样品的Shannon指数(4.701)大于对照组.较高的物种多样性有利于维持微生物群落结构的稳定,微生物群落结构的变化主要源于ZVI的添加.在添加ZVI-AC的系统中,梭菌属(Clostridium sensu sticto)成为主要的微生物,比对照组高33%.在另一项研究中,Yang等[91]研究发现添加400 mg/L Fe0NPS的实验组中狭义梭菌属(菌株Clostridium sensu stricto1与Clostridium sensu stricto3)、肠杆菌属(Enterobacter sp.)分别占66.5%、20%,而对照组狭义梭菌属(Clostridium sensu stricto1)、肠杆菌属(Enterobacter sp.)和微杆菌属(Exiguobacterium sp.)分别占11.9%、64.4%和22.5%.微生物群落结构出现变化很可能是由于Fe0NPS使系统中的氧化还原电位降低,有利于严格厌氧菌的生长.在氢气发酵过程中,氧化还原电位是影响系统产氢性能的关键因素[137].严格厌氧菌在氧化还原电位低于-200 mV的环境下表现出较好的活性,而向系统中添加了400 mg/L Fe0NPS后,系统的氧化还原电位从-133.6 mV降低至-273.3 mV,使优势菌属从肠杆菌属变为梭菌属.虽然肠杆菌属和梭菌属都能利用多种底物(如葡萄糖、蔗糖、半纤维素和纤维素)产氢,但梭菌属的产氢能力高于肠杆菌属,这是因为肠杆菌属只能通过甲酸分解途径产氢[理论产氢量为2 mol H2/(mol葡萄糖)],而梭菌属可通过丙酮酸分解途径和NADH途径产氢[理论产氢量为4 mol H2/(mol葡萄糖)]. ...
Renewable hydrogen generation by bimetallic zero valent iron nanoparticles
Relation between addition of MNPs and H<sub>2</sub> production in dark fermentation from lignocellulose<sup>[<xref ref-type="bibr" rid="R91">91</xref>, <xref ref-type="bibr" rid="R95">95</xref>, <xref ref-type="bibr" rid="R102">102</xref>-<xref ref-type="bibr" rid="R104">104</xref>, <xref ref-type="bibr" rid="R115">115</xref>]</sup>Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... , 95, 102-104, 115]Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
Biohydrogen production from sugarcane bagasse hydrolysate: Effects of pH, S/X, Fe2+, and magnetite nanoparticles
1
2017
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Dual production of hydrogen and biochar from industrial effluent containing phenolic compounds
1
2021
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Nanoparticle-associated single step hydrogen fermentation for the conversion of starch potato waste biomass by thermophilic Parageobacillusthermoglucosidasius
1
2021
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Synergistic enhancement of biohydrogen production from grass fermentation using biochar combined with zero-valent iron nanoparticles
3
2019
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
... 目前针对暗发酵产氢过程中微生物群落变化的研究主要是通过高通量测序观察微生物的相对丰度变化.Yang等[99]研究了铁纳米颗粒添加对暗发酵微生物群落的影响,发现相较于对照组,添加Fe0NPS、carbon/Fe0NPs样品的Simpson指数显著降低,Shannon指数显著升高,表明这两种添加剂显著提高了系统中微生物的多样性.此外,还可使肠杆菌属(Enterobacter sp.)和梭菌属(Clostridium sp.)的相对丰度存在显著差异.对照组、Fe0NPS和carbon/Fe0NPs的实验组中Enterobacter sp.分别占87.8%、52.8%和15.6%,Clostridium sp.分别占7.2%、40.3%和77.5%.Zhang等[100]研究了零价铁粉末-活性炭(ZVI-AC)强化生物制氢过程中微生物群落的变化,发现添加了ZVI-AC样品的Shannon指数(4.701)大于对照组.较高的物种多样性有利于维持微生物群落结构的稳定,微生物群落结构的变化主要源于ZVI的添加.在添加ZVI-AC的系统中,梭菌属(Clostridium sensu sticto)成为主要的微生物,比对照组高33%.在另一项研究中,Yang等[91]研究发现添加400 mg/L Fe0NPS的实验组中狭义梭菌属(菌株Clostridium sensu stricto1与Clostridium sensu stricto3)、肠杆菌属(Enterobacter sp.)分别占66.5%、20%,而对照组狭义梭菌属(Clostridium sensu stricto1)、肠杆菌属(Enterobacter sp.)和微杆菌属(Exiguobacterium sp.)分别占11.9%、64.4%和22.5%.微生物群落结构出现变化很可能是由于Fe0NPS使系统中的氧化还原电位降低,有利于严格厌氧菌的生长.在氢气发酵过程中,氧化还原电位是影响系统产氢性能的关键因素[137].严格厌氧菌在氧化还原电位低于-200 mV的环境下表现出较好的活性,而向系统中添加了400 mg/L Fe0NPS后,系统的氧化还原电位从-133.6 mV降低至-273.3 mV,使优势菌属从肠杆菌属变为梭菌属.虽然肠杆菌属和梭菌属都能利用多种底物(如葡萄糖、蔗糖、半纤维素和纤维素)产氢,但梭菌属的产氢能力高于肠杆菌属,这是因为肠杆菌属只能通过甲酸分解途径产氢[理论产氢量为2 mol H2/(mol葡萄糖)],而梭菌属可通过丙酮酸分解途径和NADH途径产氢[理论产氢量为4 mol H2/(mol葡萄糖)]. ...
Enhanced dark fermentative hydrogen production by zero-valent iron activated carbon micro-electrolysis
2
2015
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
... 目前针对暗发酵产氢过程中微生物群落变化的研究主要是通过高通量测序观察微生物的相对丰度变化.Yang等[99]研究了铁纳米颗粒添加对暗发酵微生物群落的影响,发现相较于对照组,添加Fe0NPS、carbon/Fe0NPs样品的Simpson指数显著降低,Shannon指数显著升高,表明这两种添加剂显著提高了系统中微生物的多样性.此外,还可使肠杆菌属(Enterobacter sp.)和梭菌属(Clostridium sp.)的相对丰度存在显著差异.对照组、Fe0NPS和carbon/Fe0NPs的实验组中Enterobacter sp.分别占87.8%、52.8%和15.6%,Clostridium sp.分别占7.2%、40.3%和77.5%.Zhang等[100]研究了零价铁粉末-活性炭(ZVI-AC)强化生物制氢过程中微生物群落的变化,发现添加了ZVI-AC样品的Shannon指数(4.701)大于对照组.较高的物种多样性有利于维持微生物群落结构的稳定,微生物群落结构的变化主要源于ZVI的添加.在添加ZVI-AC的系统中,梭菌属(Clostridium sensu sticto)成为主要的微生物,比对照组高33%.在另一项研究中,Yang等[91]研究发现添加400 mg/L Fe0NPS的实验组中狭义梭菌属(菌株Clostridium sensu stricto1与Clostridium sensu stricto3)、肠杆菌属(Enterobacter sp.)分别占66.5%、20%,而对照组狭义梭菌属(Clostridium sensu stricto1)、肠杆菌属(Enterobacter sp.)和微杆菌属(Exiguobacterium sp.)分别占11.9%、64.4%和22.5%.微生物群落结构出现变化很可能是由于Fe0NPS使系统中的氧化还原电位降低,有利于严格厌氧菌的生长.在氢气发酵过程中,氧化还原电位是影响系统产氢性能的关键因素[137].严格厌氧菌在氧化还原电位低于-200 mV的环境下表现出较好的活性,而向系统中添加了400 mg/L Fe0NPS后,系统的氧化还原电位从-133.6 mV降低至-273.3 mV,使优势菌属从肠杆菌属变为梭菌属.虽然肠杆菌属和梭菌属都能利用多种底物(如葡萄糖、蔗糖、半纤维素和纤维素)产氢,但梭菌属的产氢能力高于肠杆菌属,这是因为肠杆菌属只能通过甲酸分解途径产氢[理论产氢量为2 mol H2/(mol葡萄糖)],而梭菌属可通过丙酮酸分解途径和NADH途径产氢[理论产氢量为4 mol H2/(mol葡萄糖)]. ...
Explore the possible effect of TiO2 and magnetic hematite nanoparticle addition on biohydrogen production by Clostridium pasteurianum based on gene expression measurements
1
2016
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Ferric oxide/carbon nanoparticles enhanced bio-hydrogen production from glucose
4
2018
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
... , 102-104, 115]Relation between addition of MNPs and H<sub>2</sub> production in dark fermentation from lignocellulose<sup>[<xref ref-type="bibr" rid="R91">91</xref>, <xref ref-type="bibr" rid="R95">95</xref>, <xref ref-type="bibr" rid="R102">102</xref>-<xref ref-type="bibr" rid="R104">104</xref>, <xref ref-type="bibr" rid="R115">115</xref>]</sup>Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... , 102-104, 115]Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
Ferric oxide/date seed activated carbon nanocomposites mediated dark fermentation of date fruit wastes for enriched biohydrogen production
2
2021
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
Relation between addition of MNPs and H<sub>2</sub> production in dark fermentation from lignocellulose<sup>[<xref ref-type="bibr" rid="R91">91</xref>, <xref ref-type="bibr" rid="R95">95</xref>, <xref ref-type="bibr" rid="R102">102</xref>-<xref ref-type="bibr" rid="R104">104</xref>, <xref ref-type="bibr" rid="R115">115</xref>]</sup>Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... -104, 115]Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
Optimisation and enhancement of biohydrogen production using nickel nanoparticles — a novel approach
2
2013
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
... Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
Augmented biohydrogen production from rice mill wastewater through nano-metal oxides assisted dark fermentation
1
2021
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Improving effect of metal and oxide nanoparticles encapsulated in porous silica on fermentative biohydrogen production by Clostridium butyricum
2
2013
... 金属纳米颗粒通过提高氢化酶的活性促进暗发酵产氢过程已经得到界内的认可,一些学者近年来利用铁、镍和其他金属纳米颗粒及其掺杂复合材料等来促进暗发酵产氢的研究总结见表2.Han等[60]研究了不同浓度的Fe2O3NPs对以丁酸梭菌为优势菌群的混合厌氧细菌培养系统在以蔗糖为底物的间歇反应器中介导的暗发酵生物制氢的影响,发现在最适温度35℃,pH=8.48,纳米颗粒的浓度为200 mg/L的条件下,最大产氢量达3.21 mol H2/(mol蔗糖),比对照组(添加量为0)的产氢量高32.64%.不同金属纳米颗粒的复合材料也有相应的研究,Beckers等[110]研究了包裹在多孔SiO2中的Fe x O y 、Pd、Ag、Cu复合纳米颗粒(FeNPs@SiO2,PdNPs@SiO2,AgNPs@SiO2,CuNPs@SiO2)对暗发酵厌氧丁酸梭菌产氢的影响,发现与对照组(不添加纳米颗粒)相比,复合纳米颗粒均对产氢有促进作用,其中FeNPs@SiO2纳米颗粒的作用最明显,氢气总产量提高了38%,平均比产氢速率从44.7 ml/h提高到61.8 ml/h. ...
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Enhanced fermentative hydrogen production from industrial wastewater using mixed culture bacteria incorporated with iron, nickel, and zinc-based nanoparticles
2
2019
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
Comparative evaluation of fermentative hydrogen production using Enterobacter cloacae and mixed culture: effect of Pd (Ⅱ) ion and phytogenic palladium nanoparticles
1
2014
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Enhancement effect of silver nanoparticles on fermentative biohydrogen production using mixed bacteria
1
2013
... Application of different MNPs in dark fermentative hydrogen productionTable 2
序号
微生物种类
基质
条件参数
纳米材料
H2产量提高效果①
1
Enterobacter
straw
pH 7,37℃
Fe0NPs
提高73.1%[91]
2
Enterobacter cloacae DH-89
glucose
pH 7,37℃
FeNPs
提高230%[94]
3
Clostridium pasteurianum
glucose
35℃
Fe2O3 NPs
提高52.5%[80]
4
Clostridium butyricum
sucrose
pH 7,35℃
α-Fe2O3 NPs
提高32.64%[60]
5
Enterobacter aerogenes
glucose
pH 6.0,37℃
γ- Fe2O3 NPs
提高17%[24]
6
mixed culture bacteria
inorganic salt
pH 6,37℃
iron oxide NPs
提高81.4%[95]
7
heat pretreated sludge
sugarcane bagasse
pH 5.0,30℃
Fe3O4 NPs
提高69.6%[96]
8
anaerobic sludge
paper mill waste water
pH 7.5
Fe3O4 NPs
提高127.4%[97]
9
Parageobacillus thermoglucosidasius KCTC 33548
glucose, starch
pH 6.5,55℃
Fe3O4 NPs
提高315%[98]
10
heat pretreated sludge
grass
pH 7,37℃
Fe0 NPs / biochar
提高89.8%[99]
11
anaerobic bacteria
glucose
pH 7,30℃
Fe0 NPs / activated carbon
提高50.2%[100]
12
Enterobacter aerogenes
glucose
pH 6.8,37℃
Fe0 NPs /chitosan
提高30%[29]
13
Clostridium pasteurianum
glucose
pH 7,35℃
α-Fe2O3&TiO2 NPs
提高24.9%[101]
14
anaerobic mixed bacteria
glucose
pH 7,37℃
Fe2O3-Fe3O4 NPs /carbon
提高33.7%[102]
15
Enterobacter aerogenes
fruit waste
pH 6.5,37℃
Fe3O4 NPs /DSAC
提高204.5%[103]
16
mixed culture bacteria
gelatinaceous wastewater
pH 6,35℃
Fe3O4/graphene oxide
提高41.9%[104]
17
Enterobacter aerogenes
glucose
pH 6,37℃
ferric citrate NPs
提高50.45%[105]
18
mesophilic bacteria
starch
pH 5.0~6.0,37℃
Fe0 NPs, Ni0 NPs
提高37%[2]
19
Clostridium butyricum
glucose, starch
pH 6.8,37℃
Fe0 NPs&Ni0 NPs
提高28%[106]
20
hydrogen-producing bacteria
anaerobic sludge
pH 5.0,37℃
Fe2O3 NPs, NiO NPs
分别提高24%和16%[107]
21
thermophilic mixed bacteria
glucose
pH 5.5,60℃
α-Fe2O3 NPs, NiO NPs
分别提高34.38%和5.47%[4]
22
mixed consortia
glucose
pH 5.6,35℃
Ni NPs
提高22.71%[108]
23
Clostridium butyricum
glucose
pH6.9,37℃,55℃
NiFe2O4 NPs
分别提高38.6%(37℃),28.3%(55℃)[30]
24
Bacillus anthracis
palm oil
pH 7,37℃
NiO NPs, CoO NPs
分别提高151%和167%[21]
25
Clostridium beijerinckii
rice mill wastewater
pH 7,37℃
NiO NPs, CoO NPs
分别提高109%和90.4%[109]
26
Clostridiumbutyricum
glucose
pH 7.6,30℃
Fe NPs@SiO2, Pd NPs@SiO2, Ag NPs@SiO2, Cu NPs/SiO2
提高38%[110]
27
mixed consortia
glucose
pH 5.5,50℃
ZnO NPs
提高29%[111]
28
Clostridiumbutyricum
sucrose
pH 7.2,35℃
Au NPs
提高61.7%[38]
29
Enterobacter cloacae
glucose
pH 7,37℃
Pd(Ⅱ) NPs
在单一菌种和混合菌种培养条件下分别提高1.5%和9%[112]
30
mixed culture dominated by Clostridium species
glucose
pH 8.0~9.4,35℃
Ag NPs
提高67.3%[113]
① 与不添加纳米材料的对照组比较. ...
Stress response of pseudomonas species to silver nanoparticles at the molecular level
Role of L-cysteine and iron oxide nanoparticle in affecting hydrogen yield potential and electronic distribution in biohydrogen production from dark fermentation effluents by photo-fermentation
Relation between addition of MNPs and H<sub>2</sub> production in dark fermentation from lignocellulose<sup>[<xref ref-type="bibr" rid="R91">91</xref>, <xref ref-type="bibr" rid="R95">95</xref>, <xref ref-type="bibr" rid="R102">102</xref>-<xref ref-type="bibr" rid="R104">104</xref>, <xref ref-type="bibr" rid="R115">115</xref>]</sup>Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
... , 115]Fig.7
Mullai等[108]研究镍纳米颗粒对暗发酵产氢的影响,发现在pH=5.6、温度30~35℃时,产氢量可提高21%,且镍纳米颗粒浓度为5.67 mg/L时,在最佳条件下可获得4400 ml的累积产氢量.Yang等[91]研究发现Fe0NPs可使产氢量和比产氢速率分别提高73.1%和128.3%,这是由于Fe0NPs加快了铁氧还蛋白和氢化酶之间的电子转移速率,从而增强了微生物的活性.Zhang等[30]比较了中温(37℃)和高温(55℃)下添加不同剂量的NiFe2O4NPs对微生物产氢的效果.37℃时添加量为100 mg/L和55℃时添加量为200 mg/L时,得到最高的H2产率分别为222 ml H2/(g葡糖糖)和130 ml H2/(g葡糖糖),较对照组分别提高38.6%和28.3%.这是由于NiFe2O4NPs经微生物的内吞作用释放的铁和镍有利于铁氧还蛋白和氢化酶的合成,增强了丁酸盐产氢途径,优化了微生物群落结构,增加了丁酸梭菌的丰度. ...
Enhanced biohydrogen production from date seeds by Clostridium thermocellum ATCC 27405
Revealing the mechanisms of alkali-based magnetic nanosheets enhanced hydrogen production from dark fermentation: comparison between mesophilic and thermophilic conditions
Improving hydrogen and methane co-generation in cascading dark fermentation and anaerobic digestion: the effect of magnetite nanoparticles on microbial electron transfer and syntrophism
MNPs involve in intracellular and extracellular electron transportation for H<sub>2</sub> and CH<sub>4</sub> production <i>via</i> IET interaction<sup>[<xref ref-type="bibr" rid="R120">120</xref>]</sup>Fig.8
TEM images of MNPs stuck to the bacterial surfaces<sup>[<xref ref-type="bibr" rid="R60">60</xref>, <xref ref-type="bibr" rid="R119">119</xref>]</sup>Fig.9
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
... [120]MNPs involve in intracellular and extracellular electron transportation for H<sub>2</sub> and CH<sub>4</sub> production <i>via</i> IET interaction<sup>[<xref ref-type="bibr" rid="R120">120</xref>]</sup>Fig.8
TEM images of MNPs stuck to the bacterial surfaces<sup>[<xref ref-type="bibr" rid="R60">60</xref>, <xref ref-type="bibr" rid="R119">119</xref>]</sup>Fig.9
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
TEM images of MNPs stuck to the bacterial surfaces<sup>[<xref ref-type="bibr" rid="R60">60</xref>, <xref ref-type="bibr" rid="R119">119</xref>]</sup>Fig.9
SEM images of bacterial nanowire produced by response of surface protein of <i>E. aerogenes</i> cells and Fe<sub>2</sub>O<sub>3</sub>NPs<sup>[<xref ref-type="bibr" rid="R24">24</xref>]</sup>Fig.10
Syntrophy and interspecies electron transfer in methanogenic microbial communities
1
2020
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
Microbial symbiosis: a network towards biomethanation
1
2020
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
A new model for electron flow during anaerobic digestion: direct interspecies electron transfer to Methanosaeta for the reduction of carbon dioxide to methane
1
2014
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
Methanogenesis facilitated by electric syntrophy via (semi)conductive iron-oxide minerals
1
2012
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
Magnetite compensates for the lack of a pilin-associated c-type cytochrome in extracellular electron exchange
1
2015
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
Electrical transport along bacterial nanowires from Shewanella oneidensis MR-1
1
2010
... 具有导电性的磁铁矿或金属铁氧化物具有促进种间电子传递(interspecies electron transfer, IET)的功能,IET又可分为介导型种间电子传递(MIET)和种间直接电子传递(DIET)[120].在MIET机制中,电子通过可溶性介质(如甲酸盐、H2)传递[121-122],而DIET是一种更为高效的种间电子传递方式,微生物细胞通过细胞膜表面的导体(如纤毛)进行电子传递,不需要介质的参与[123].金属纳米颗粒可通过聚集形成微生物细胞间的“非生物纳米导线”,弥补如细胞色素C、纤毛等生物导体的不足,促进种间直接电子传递和细胞膜内电子传递(图8)[120, 124-125].Han等[60]通过透射电镜观察到了暗发酵制氢过程中Fe2O3NPs黏附在混合菌群细胞表面[图9(a)],这可能有助于细胞的固定,并增加其与基质接触的机会.此外,在暗发酵初期,混合菌群中的某些细菌可通过胞外呼吸的方式,将细胞内氧化有机物产生的电子跨膜传递到胞外电子受体表面,而附着在细菌表面的Fe2O3NPs可作为胞外呼吸的末端电子受体.并且由于部分铁氧化物具有良好的导电性,在胞外电子传递过程中可充当导电材料.异化铁还原菌是可进行胞外呼吸的典型代表微生物,能以含三价铁的铁氧化物作为外源末端电子受体,将三价铁还原为二价铁.Zhang等[30]在混合菌群暗发酵产氢过程中投加NiFe2O4NPs,发现虽然NiFe2O4NPs中铁元素多为三价铁形式,但系统液相中铁却以Fe2+形式存在,这表明NiFe2O4NPs中的铁可被异化铁还原菌作为电子受体利用,梭菌作为一种典型的发酵型异化铁还原菌在以葡萄糖为碳源的铁还原环境中起主导作用.金属纳米颗粒作为介导电子供体菌与电子受体菌之间的“桥梁”在种间电子传递过程中发挥作用.Lin等[24]通过扫描电镜观察发现,与未添加Fe2O3NPs的细胞相比,添加Fe2O3NPs的肠杆菌细胞表面出现更多的团聚体,这些团聚体是细胞表面蛋白与Fe2O3NPs响应产生的生物纳米导线(bacterial nanowire,BN)(图10),能促进葡萄糖进入细胞,同时也在细胞间的电子传递中起到重要作用.El-Naggar等[126]也证实了电子会沿着西瓦氏菌的生物纳米导线传递. ...
Mitigation of CuO nanoparticle-induced bacterial membrane damage by dissolved organic matter
High yield expression of an AHL-lactonase from Bacillus sp. B546 in Pichia pastoris and its application to reduce Aeromonas hydrophila mortality in aquaculture




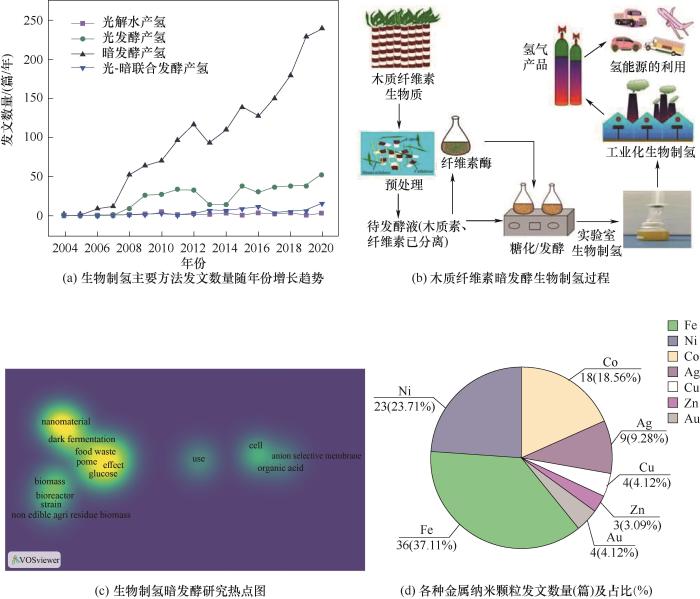
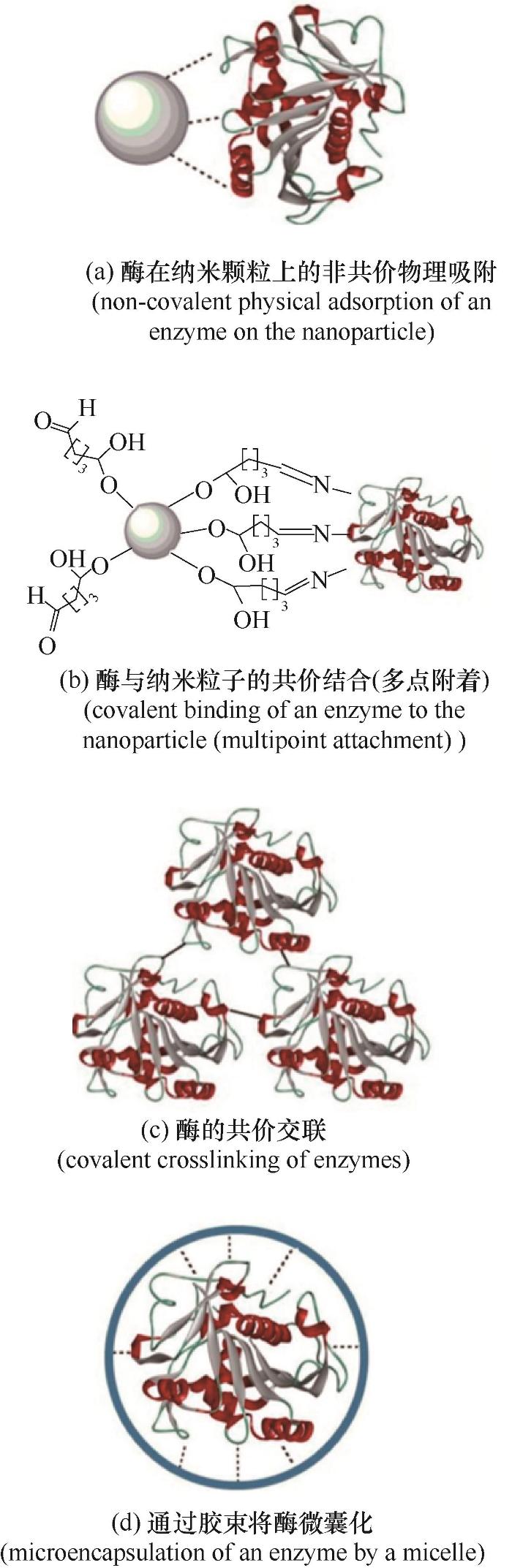
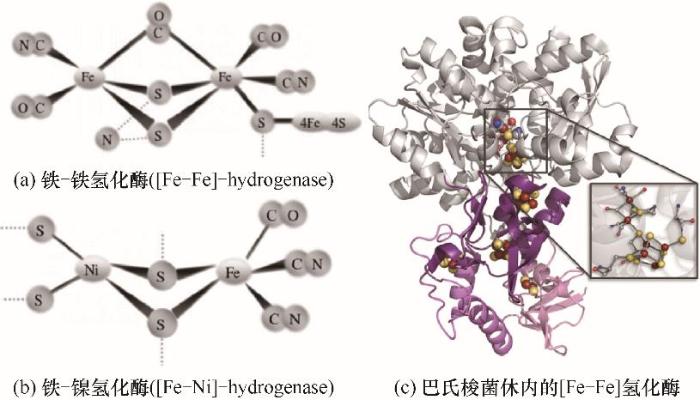
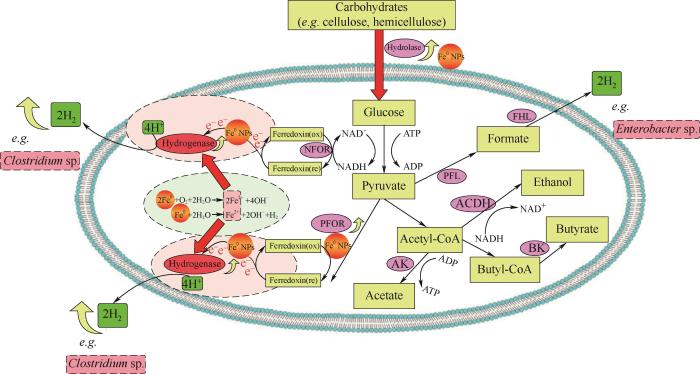
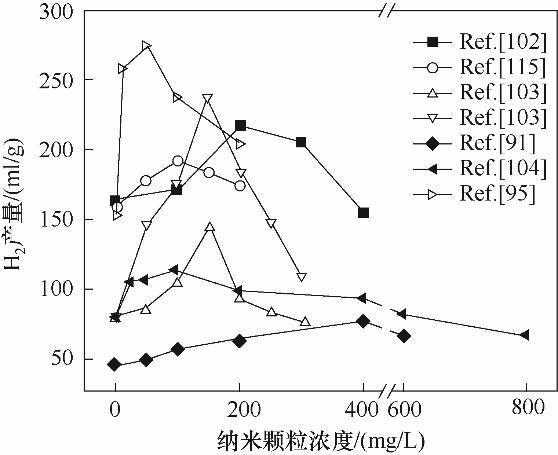
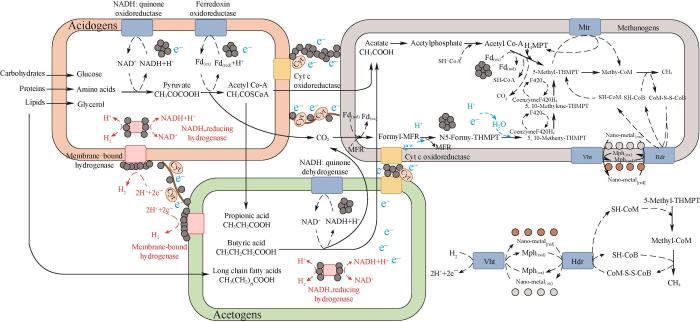



 京公网安备 11010102001995号
京公网安备 11010102001995号{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}